Pourquoi le chromosome Y dégénère-t-il ?
Chez de nombreuses espèces, les individus mâles possèdent un chromosome Y qui ne recombine plus avec le X et ne contient qu'un petit nombre de gènes. La principale théorie proposée depuis plus de 40 ans postule que cette dégénérescence est causée par l’inefficacité de la sélection naturelle en absence de recombinaison. L’étude parue dans Current Biology, propose une nouvelle théorie basée sur l’instabilité des régulateurs génétiques, montrant que la dégénérescence du Y et la compensation de l’expression des gènes du X peuvent découler d'un même processus. L’article est issu de la collaboration entre le Centre d'Ecologie Fonctionnelle et Evolutive (Univ Montpellier/CNRS/Univ Paul Valéry Montpellier 3/EPHE/ IRD), l’Institut Radcliffe (Harvard University), l’unité de recherche Biologie évolutive et écologie des algues (CNRS) et la Station Biologique de Roscoff (Sorbonne Université).
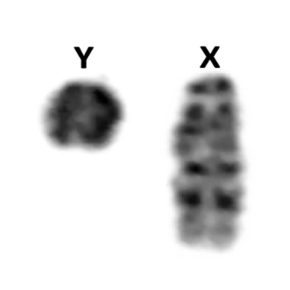
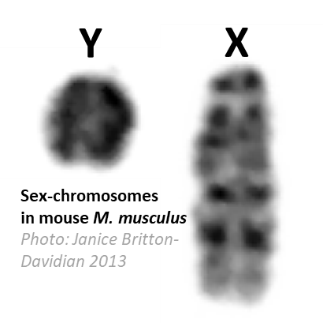
Chez de nombreuses espèces, le déterminisme du sexe est chromosomique: par exemple, chez l’homme, le mâle est XY et la femelle XX. Les chromosomes X et Y sont souvent très différents: le chromosome Y est généralement bien plus petit que le X (cf. photo) et contient très peu de gènes fonctionnels. De ce fait, cette situation s’accompagne souvent de mécanismes d’ajustement de l’expression des gènes du chromosome X pour compenser leur absence sur le Y (la compensation de dosage). Ainsi, chez les mammifères, un seul des deux chromosomes X des femelles est exprimé, permettant de conserver un taux d'expression des gènes similaire chez les mâles (qui ne possèdent qu'un X) et les femelles.
Depuis plus de 40 ans, on pense que la disparition des gènes du Y est due à un processus de « dégénérescence » correspondant à une accumulation de mutations délétères causée par l’inefficacité de la sélection naturelle en absence de recombinaison. En effet, le chromosome Y ne recombine pratiquement pas avec le X (sauf sur une petite portion, la région pseudo-autosomale). Or, on sait que l'absence de recombinaison peut conduire à une accumulation progressive de mutations délétères au cours des générations: c'est le phénomène du "cliquet de Muller" (du nom de H.J. Muller, généticien américain ayant décrit ce phénomène dans les années 1930). Appliquée aux chromosomes sexuels, cette théorie se heurte cependant à plusieurs difficultés. En particulier, le cliquet de Muller est un processus relativement lent dans les populations de grande taille, et devient quasiment inopérant lorsque les régions non-recombinantes ne contiennent qu'un petit nombre de gènes. Or, l’étude fine de nombreux chromosomes sexuels a révélé que l'arrêt de la recombinaison et la dégénérescence surviennent souvent de façon progressive, par portions du Y (des « strates ») ne contenant parfois qu'assez peu de gènes. Enfin, cette théorie ne s’est jamais vraiment penchée sur l’évolution de la compensation de dosage, supposant qu’il s’agissait d’un processus indépendant ne survenant qu'après la dégénérescence.
Dans un article publié dans Current Biology, Lenormand et al. proposent une nouvelle théorie pour expliquer la dégénérescence des chromosomes Y. Pour cela, les auteurs modélisent explicitement l’évolution de l'expression des gènes présents sur les chromosomes sexuels. Après l'arrêt de la recombinaison, le taux d'expression des gènes présents sur le X et le Y est libre de diverger de façon aléatoire, tant que le niveau d'expression global est maintenu. Lorsqu'un gène présent sur le Y devient peu exprimé, celui-ci aura tendance à accumuler des mutations, menant à sa dégénérescence. A l'inverse, lorsqu'un gène du X se retrouve moins exprimé que sa copie présente sur le Y, il ne peut pas accumuler de mutations car celles-ci conduiraient à la production de protéines non-fonctionnelles chez les femelles. Ainsi, l’évolution des séquences régulatrices de l'expression des gènes suffit à expliquer la dégénérescence du Y ainsi que la compensation de dosage associée. Bien sûr, cette théorie n’exclut pas les processus liés au cliquet de Muller, mais elle propose un mécanisme supplémentaire pouvant conduire à la dégénérescence rapide de toute région non-recombinante du Y, si petite soit-elle. Cette théorie renouvelle donc en profondeur notre compréhension de l’évolution des régions non recombinantes des génomes eucaryotes.
Référence :
Thomas Lenormand, Frederic Fyon, Eric Sun, Denis Roze. 2020. Sex chromosome degeneration by regulatory evolution. Current Biology, in press.